|
GENETICS –The mechanism of inheritance
 |
Paul Smart - The illustrated encyclopedia of the Butterfly world |
|
|
Genetics is the study of the ways in which the characters of an organism are inherited by its offspring. A number of plants and animals have been studied intensively by geneticists because they have simplified mechanisms that lend themselves to scientific investigation and analysis. The genetic principles which have been discovered and understood in relatively simple systems c¹n then be used to explain the situations found in a wide spectrum of species. Butterflies have received very little attention from geneticists but there has been a fair amount of work done on moths.
In the absence of detailed information specifically relating to Butterflies it is helpful that much of our understanding of the genetic processes which has been obtained by exhaustive study of the fruit fly Drosophila is probably equally applicable to butterflies.
Every cell in the body of a butterfly, or any other organism, contains in the nucleus microscopic structures called chromosomes. These chromosomes are a kind of chemical blueprint for all the structures and functions of the animal. The number of chromosomes in each cell is normally constant for any particular species, but may differ from one species to the next. The chromosomes occur in pairs and the two members of each pair are said to be homologous. The members of each homologous pair are derived from the parents in the following way. When reproductive cells are produced the individual chromosome pairs divide into single chromosomes which each pass into different cells so that each sperm or egg has only half the full. number of chromosomes. At fertilization, sperm and egg fuse and their two half sets of chromosomes again pair up to produce a fertile egg cell that contains a full set of paired chromosomes.
The basic units of heredity are called genes and these are carried in the chromosomes. Like the chromosomes these genes are in pairs, one inherited from the father and one from the mother. In the simplest possible situation a single character is controlled by one such pair of genes. The actual situation is often more complicated because one gene pair may control several characters or alternatively several gene pairs may affect a single character. If the two genes of a pair are of similar type the animal is described as homozygous or a homozygote. Alternatively if the two genes of a pair are dissimilar then it is heterozygous and the animal is described as a heterozygote.
It should be remembered that this description refers to a particular gene pair and one animal can be both homozygous for some characters and heterozygous for others.
Knowledge of the character and behaviour of the genes is of vital importance in understanding the genetic process. The two genes in a pair do not fuse or mix even when they are dissimilar in character. They retain their integrity and when reproductive cells are produced they can separate again. The separation (tách rời) of every pair of genes into different reproductive cells is termed segregation. These basic principles of inheritance may be referred to as 'Mendelian' after Father Gregor Mendel, an Augustinian abbot, who investigated the patterns of simple inheritance and first proposed the existence of inheritance factors. At that time, the mid - 1860s, the existence of chromosomes and genes was not known, so it was not until much later, in the 20th century, that the importance of Mendel's work was properly 'appreciated.
|

|
|
|
The Common mormon Papilio polytes - Picture: Phùng Mỹ Trung |
|
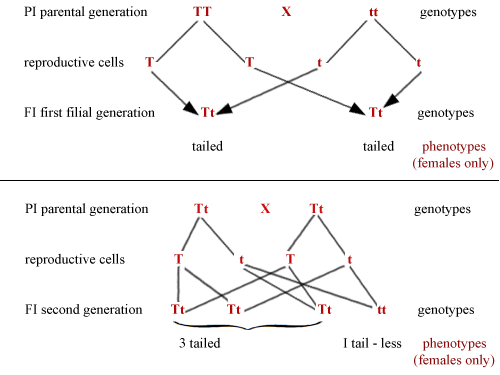
The Mendelian principle can be illustrated by the case of the oriental swallowtail butterfly Papilio memnon. The female is polymorphic; that is, it exists in more than one form (see Chapter 8) and these different forms are determined genetically. The butterflies are tailed in the form achates and tail-less in the form agenor. The tail-less butterflies are homozygous (ie have two tail-less genes) while the tailed butterflies may be homozygous (two tailed genes) or heterozygous (one tailed gene and one tail-less gene). It is the usual situation for one gene type in a pair to be dominant, suppressing the other member of the pair when they occur together in the heterozygous condition. In this species the tailed gene is dominant while the tail-less gene is recessive. It should be emphasized, however, that the recessive gene still retains its genetic potential for future generations, but it will not be expressed in the appearance of the butterfly unless it is paired with a second tail-less gene. This important point is underlined by the use of the terms 'genotype' for the genetic make-up of an individual and 'phenotype' for its actual appearance. This can be illustrated where the fate of the genes is traced through to the next generation in the following diagram T represents the dominant tailed gene (achates) while represents the recessive tail-less gene (agenor). In the top diagram parents are both homozygous.
Because T is dominant over t, it will be seen that the first generation females are all tailed in appearance, while carrying both genes. It should be remembered that the male parent may carry the genetic constitution for achates or agenor but does not show either of these forms since they are exdusively female forms.
If a tailed F1 female of this brood is paired with one of her brothers the resulting F2 females will be tailed and tail-less in the ratio 3 : 1 respectively, as in the bottom diagram. Three of the second generation have the dominant T gene, so have tails; one has no T factor (only the t gene) so has no tail. In practice this ratio may not be exactly achieved since some of these characters will be inherited by males and will not therefore be expressed in the F2 generation. This system of two alternatives, a character being either present or absent, is much more common than situations where 'mixing' occurs to give intermediate types between the parental forms.
|
|
|
Another example of this genetic process is proved by the small copper butterfly Lycaena phlaeas. The male and female of this species are similar in colour. They have coppery fore wings spotted with blackish-brown and brown hind wings with a copper band close to the hind margin. There is a rare variety called obsolela in which this copper band is missing. It has been shown that the presence of this band is determined by a simple pair of genes. The typical form of the butterfly is dominant while the obsoleta gene is recessive. It is similar to the situation found in the oriental swallowtail except that the variety may be found in both sexes. When a normal parent with two normal genes is paired with an obsoleta parent which carries two obsoleta genes the first generation are all heterozygotes and they all appear normal because they all contain the normal gene which is dominant over their other obsoleta gene. When two of these heterozygotes are paired the offspring consist of normal and obsoleta fonns in the ratio 3 : 1. This ratio is what is visibly apparent in the phenotypes, in fact the ratio is 1 : 2 : l because there is one homozygous normal, two heterozygous normal and one homozygous obsoleta form.
Further examples of departure from the normal form are described in "see Variation". Butterflies such as the silverwashed fritillary (Argynnis paphia) and the clouded yellow (Colias crocea) have dimorphic females which are produced genetically. The variant genes can only find expression in the internal developmental environment of the female of the species and they are therefore called sex-controlled genes. The examples which are given are dominant genes which produce the variety in the heterozygous animals, unlike obsoleta referred to above. Homozygous forms of these two species are unknown, presumably because they are non-viable and cannot survive in the wild.
Gynandromorphism.
Gynandromorphs are butterflies which exhibit the external characters of both the male and female in the same individual. Such specimens are sometimes incorrectly referred to as hermaphrodites.
These abnormal forms may be difficult to detect when there are no great differences in the appearance of the sexes, but in species with pronounced sexual dimorphism such insects look very odd indeed. In extreme examples the wings on one side belong to one sex while those on the opposite side belong to the other sex.
The sex of an organism is specified by its genetic make-up. In mammals the genetic composition determines whether the gonads are male (testes) or female (ovaries). All the other characters of each sex are then produced under the influence of the particular hormones which are produced by the gonads. Circulating in the blood, these hormones reach every part of the animal and so the whole body assumes a male or female character. In butterflies the situation is different since the sexual appearance of every individual part of the body is determined by the genetic composition of its component cells. This is how it is possible for different parts of a butterfly to
behave independently and have the appearance of different sexes.
|

|
|
|
The Redbase jezebel Delias pasithoe - Picture: Phùng Mỹ Trung |
|
Butterfly sex is determined by the sex chromosomes. There are two types of sex chromosomes, X and Y. A butterfly which possess two X chromosomes is male, while a butterfly which carries both X and Y is female. Sex is inherited in the same way as other characteristics. The homologous pair of sex chromosomes divides during reproduction and the male produces sperms which carry one X chromosome each. The female produces eggs with either X or Y chromosomes in equal numbers. The resulting progeny are males and females in approximately equal numbers.
The actual sex determination depends on the presence of the extra X chromosome. Some abnormal forms with an extra Y chromosome are still male because two X chromosomes are present. Gynandromorphs are produced by abnormalities in the genetic composition of the butterfly. One way in which they can be produced is if the butterfly develops from an egg with two nuclei, one containing XX chromosomes and the other containing XY. Another way is if an error occurs during primary cell division of a male embryo so that an X chromosome is lost by one of the cells. Since it is the quantity of X chromosomes that determines the sex this results in a butterfly which is half normal male (XX) and half X, which is effectively female although there is no Y chromosome.
Halved gynandromorphs are generally rare and most gynandrous forms exhibit an unequal mixture 'of male and female characters. The appearance of the specimen of the adonis blue Lysandra bellargus may be due to another error during cell division. In this instance the insect is predominantly female. It is possible that during a late stage in wing development during division of the chromosomes the X chromosomes may have failed to separate. This would result in some cells having an XXY genetic make-up and others with Y alone. The latter would certainly die, while the cells with XXY would be male in character because of the two X chromosomes. These would develop male coloration giving rise to an insect with a mosaic appearance. Sometimes only a small part of the insect will be affected, as in the case of the female orange tip Anthocharis cardamines which shows a small patch of the orange scaling which is exclusively a male character
Intersexuality
Intersexes are mixed forms which originate in a different way from gynandromorphs. To understand how they arise it is necessary to understand the way in which sex is determined by the X chromosomes. It is the quantity of X chromosome material, or its relative strength, which determines the sex. One X chromosome is sufficient only to produce a female, but when there are two X chromosomes there is sufficient X influence to produce a male. Sex is not determined by any particular agent carried in the chromosome. This is realized when it is appreciated that a male whose sex is determined by having two X chromosomes inherited one from his mother and that single X chromosome was what determined his mother's female sex. Similarly if he produces female offspring their sex will be determined by one of his X chromosomes. This works effectively provided that the 'dose' of sex factor carried by an X chromosome is always the same. Problems arise when individuals from widely separate races of the same species mate and produce offspring. Where races have been isolated and are unable to meet normally in the wild they may have developed X chromosomes of different 'strength. A single X chromosome from race A may be equivalent in terms of the dose of sex factor to two X chromosomes from race B. In such an instance crossing individuals from separate races may result in offspring which develop as intersexes, or a peculiar mixture of the sexes, developing first as one sex and then later changing over to the other. This situation is known to occur in certain moths and may well be found in butterflies too.
It is remotely possible that the blue females found in Morpho aega are produced by genetic imbalance and are intersexual in origin. The difference between the colours of these and normal females is structural (see Coloration) and is not due to pigments. The form Pseudocypis (shown here) demonstrates scales of a type generally found in males of this species. Females with restricted areas of blue scaling are frequently met with, and these 'blue' forms are said to be much more frequent in some populations 'than others.
|


